Wednesday, August 12, 2015
Wednesday, July 29, 2015
Diseases with an upside!
Diseases with an upside.
By Rich Feldenberg

Since life’s earliest emergence on planet earth, disease has been our constant and unwelcome companion. Even the first single celled organisms were susceptible to break down, nutritional deficiencies, and harmful genetic mutation. When single celled life upgraded to the multicellular stage, finally becoming large, it was then susceptible to a host of new disorders, such as cancer that interfered with the organization and growth of cells that now had to survive as part of a collective. Humankind is no different than the rest of the animal kingdom in this regard, and throughout human existence disease has lead to untold suffering, death, and at times the threat of total extinction. It may therefore be surprising to learn that some diseases confer protection against other types of illness, and this seems to account for the high prevalence of some of these disorders in the human population. If the protective benefit of the disease mutation on a large portion of the population outweighs the suffering and death of a small portion of the population, natural selection will swing the balance in favor of keeping those mutations in the gene pool. Not only may the disease mutation simply persist in the gene pool, but it may become very prevalent because it is selected for in the right environment, where the other illness it protects against is a major threat. To illustrate how this works I’ll give some detail on two well known examples of diseases and their upside - in other words, how they protect against other threats to our species.
The first example is that Sickle Cell Anemia (SCA), which has the best documented evidence as to its evolutionary risk versus benefit ratio in its effected population. Sickle Cell Anemia is a genetic disease that causes anemia (low red blood cell counts), and can lead to painful, and potentially deadly pain crises. It is inherited as an autosomal recessive trait - meaning that if you receive one copy of the mutated gene from each of your parents, then you have two abnormal copies of the gene (are homozygous, in the language of genetics) and will have the disease. Each of your parents, however, has only one mutant copy and also one normal copy (is heterozygous), and so is only a carrier (has sickle cell trait) and will not show symptoms of the disease under normal circumstances.
SCA is due to a single base switch in the DNA that codes for the beta-chain of the hemoglobin molecule. Adult hemoglobin is made of two alpha chains and two beta chains. This is the major oxygen carrying protein in the blood, although, there are other versions of hemoglobin that are produced (one example is fetal hemoglobin with two alpha chains and two gamma chains). In SCA, there is a substitution of the amino acid glutamic acid for valine at the 6th amino acid in the beta chain. Since valine is more hydrophobic than glutamic acid this has the unfortunate consequence of causing the hemoglobin molecules to polymerized and compact together, deforming the shape of the red blood cells (RBCs) that carry them, into a sickle shape - hence the name Sickle Cell Anemia. The polymerization event is more likely to happen if the affected individual is dehydrated, in a low oxygen state (hypoxic), or otherwise ill with another illness. The deformed red blood cells can not get through the tiny capillaries very well, causing blockages that deprive tissues of blood and oxygen. The result is pain and organ damage.

Over time, people with SCA damage their spleen so badly that they lose the its important immune function, which normally you against encapsulated bacterial infections. These are certain bacteria that are surrounded by a polysaccaride capsule, that helps them to escape detection by the immune system. Someone without a functioning spleen can then die of these types of infections, whereas those with normal spleens would be able to fight off the infection easily. The blockages to blood flow due the abnormal sickle shaped RBCs can lead to strokes and to Acute Chest Syndrome. If people with SCA become infected with the common virus Parvovirus B19, they can develop severe life threatening anemia, with hemoglobin levels that get so low they can develop heart failure.
Sickle cell anemia is common in sub-Saharan Africa, and about 300,000 are born with disease each year. All the complications of SCA listed above can be fatal so why would this disorder have such a high prevalence? The answer seems to be that although people with full blown Sickle Cell Anemia are at a most definite disadvantage from a survival aspect, those who are carriers of SCA are protected against another common killer - Malaria. Malaria is an infectious disease caused by the protozoan Plasmodium. It has a complex life cycle, part of which is spent inside the mosquito Anopheles, and part is spent inside a vertebrate host - such as a human. When an infected female mosquito bites a human, the organism is transmitted into the persons blood stream where it travels to the liver, infects liver cells, reproduces, and then is released back to the bloodstream where it infects RBCs. The symptoms of Malaria include fever, vomiting, joint and muscle pain, headache, and in some cases seizures. As the Plasmodium organism goes through its life-cycle within the host, from liver to RBC and back again (these are known as the liver phase and the erythrocytic phase respectively), the symptoms return in a cyclical fashion. In some cases the organism passes through the blood-brain barrier leading to Cerebral Malaria, which is a very serious complication. Malaria has a high mortality rate if untreated - as would have been the case before the age of modern medicine.

It was observed, early on, that in regions endemic to malaria, people who were carriers of the sickle cell mutation showed resistance to the malaria infection, and that full blown SCA has a high prevalence in those same regions where malaria is endemic. Further studies confirmed that those individuals who are carries for the sickle cell mutation, do in fact, enjoy a protection due to their gene mutation. Unfortunately, those with actual sickle cell anemia (homozygous for the gene mutation) are not protected against malaria. Not only do they have to suffer the fate of SCA, but if they get malaria they have a worse prognosis because the malaria damages their already vulnerable RBCs.

For a long time it was thought that sickle cell trait most likely confers its malarial protection by making it difficult for Plasmodium organisms to infect the abnormally shaped RBCs, and that the abnormal RBCs are removed more readily by circulating macrophages, helping to rid Plasmodium infected cells more readily. More recent research seems to suggest that the protective mechanism is more complex that that, and involves the up regulation of an enzyme called heme oxygenase-1(HO-1). HO-1 causes the breakdown of heme, and the release of carbon monoxide (CO), iron, and biliverdin, resulting in an anti-inflammatory effect. HO-1 is upregulated or produced to a greater extent in RBCs that have the abnormal hemoglobin associated with SCA, and it is the production of CO that seems to have a detrimental effect for the Plasmodium organisms. It confers protection against cerebral malaria, and decreased mortality for those with sickle cell trait who become infected with malaria. This might also be the answer to why several other diseases or disease traits have also been observed to offer protection against malaria, such as thalassemia trait and Glucose-6-Phosphate Deficiency. These disorders might also increase the activity of HO-1.
We’ll move now to another deadly disease that seems to have remained in the population because it offered a survival advantage. This is the kidney disease called Focal Segmental Glomulosclerosis (a real mouthful) or just plain old FSGS for short. FSGS can be caused by chronic infections, such as hepatitis or HIV, but many cases are due to a genetic mutation. It is a subset of the genetic form that may have been selected for to protect against Sleeping Sickness. In FSGS the tiny filters in the kidneys, called glomeruli, become scarred until they can no longer filter. This can eventually progress to kidney failure and the need for dialysis or kidney transplant. Kidney failure is fatal without modern medical care and FSGS is one of the more common causes for young people to be on dialysis. Its also, often more common and resistant to therapy in African Americans and other people of African descent.
Some people with the genetic form of FSGS have a mutation in a gene called APOL1, and if you are an individual with two mutated copies of the APOL1 gene, your risk of developing FSGS and kidney failure is 17 times higher than if you have two normal copies of the gene. That adds up to around a 4% chance of developing FSGS over your lifetime if you are homozygous for mutant APOL1. This mutation is also thought to explain 18% of all cases of FSGS that currently exist. There are two types of mutations in the APOL1 gene that can increase risk for FSGS kidney disease. These is the G1 variant, which contains two amino acid substitutions - one is a replacement of glycine for serine at amino acid 342 in the protein (S342G), and the other switch is a replacement of methionine for isoleucine at amino acid 384 in the protein (I384M). You have to have both of these switches you have the G1 variant. The other variant is the G2 variant where 6 base pairs are deleted in the DNA coding for APOL1 starting at base 388. People can have either a G1 variant or a G2 variant, but never have both types.
APOL1 is a protein that circulates in the blood and is part of the high-density lipoprotein (HDL - otherwise known as the “good” cholesterol). Exactly how the mutated form of APOL1 causes kidney disease is still not known. What is known, however, is that those individuals with either a G1 or G2 specific gene mutation in APOL1 have protection against African Sleeping Sickness, caused by the protozoan Trypanosoma brucei. This tiny single celled eukaryotic organism is transmitted to its human host by the bite of the tsetse fly. It is a common and dangerous disease in sub-Saharan Africa. In 1990 it caused 34,000 deaths, but the death rate dropped to 9000 in 2010, thanks to efforts of the World Health Organization to prevent and treat the infection.
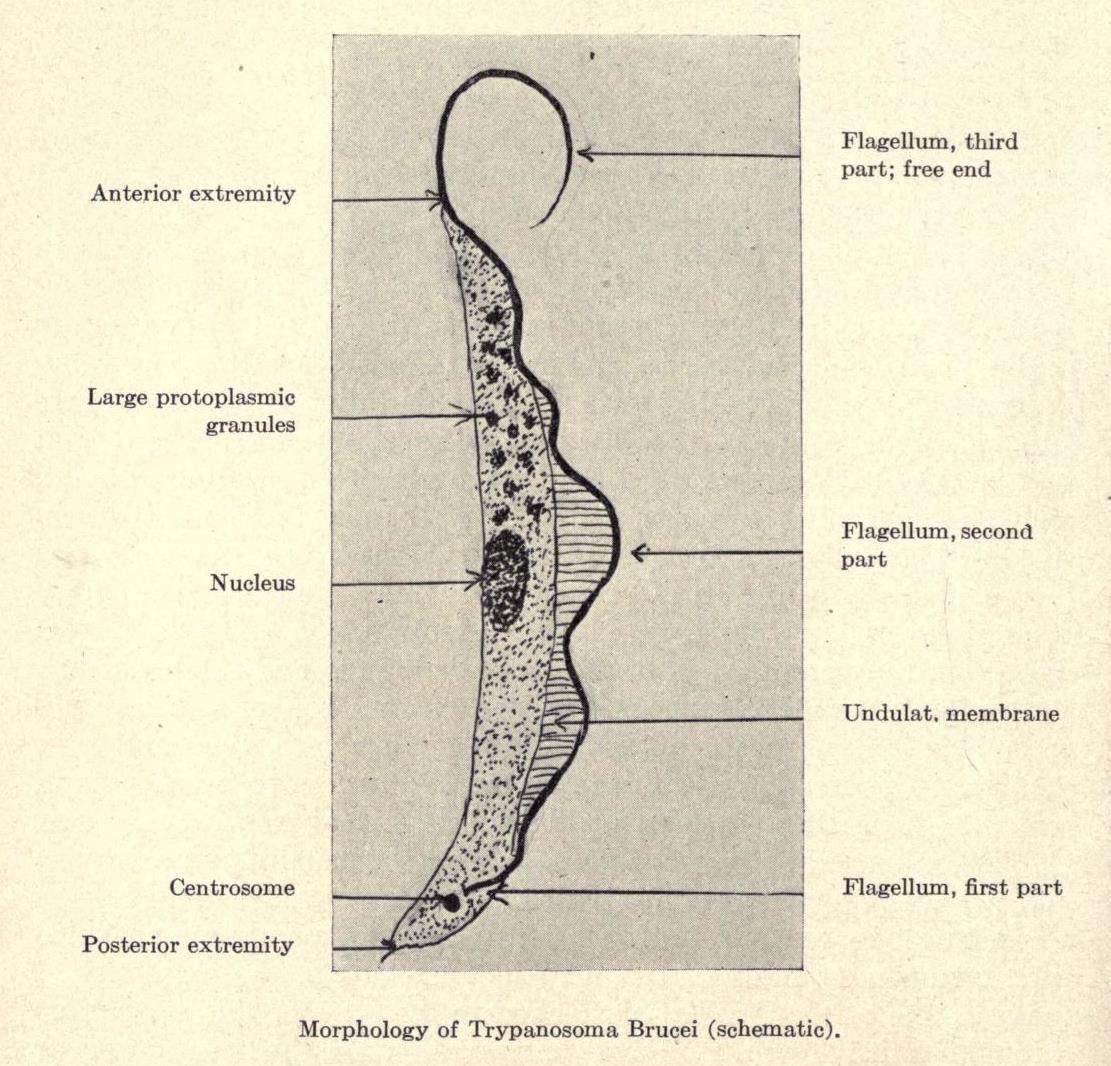
Those affected by the parasite experience two distinct stages of infection. In the first stage the victim develops headaches, fever, and severe itching. This resolves only to eventually progress to the second stage of the disease which effects the central nervous system causing confusion, paralysis, neuromuscular weakness, and sometimes psychiatric illness. There is a reversal of the sleep-wake cycle, giving the disorder its common name. Infected persons often sleep in the day and remain awake at night. Without treatment the disease always ends in the death of its victim. It can be treated with the drug pentamidine, when in the first stage, or drugs such as eflornithine or melarsoprol for second stage disease.

Like the association of Sickle Cell Anemia and malaria, those geographic regions with a high incidence of sleeping sickness also have a high incidence in the population of APOL1 G1 or G2 variants. This is because those gene variants protect against the ravages of the Trypanosomes. The APOL1 variants cause the lysis (breaking apart of the cell membrane) of Trypanosomes that cause sleeping sickness. The normal gene for APOL1 gives us resistance to other species of Trypanosomes that do infect other mammals, but are unable to harm us. The sleeping sickness Trypanosome (Trypanosome brucei rhodesiense) is immune to the normal APOL1 since it has evolved a serum resistance-associated protein (SRA) that blocks a portion of the APOL1 protein, neutralizing its anti-trypanosomal action. Not so for the APOL1 variants G1 or G2, however. They are able to get around this SRA and destroy the parasite. From an evolutionary point of view, the advantage of being more resistant to sleeping sickness in an area of high risk, outweighs the cost of having a higher than average chance of kidney disease. There is no advantage, however, to having these variants if your ancestors originated where sleeping sickness is not a problem, so other populations aren’t found to have these gene mutations.
The two examples of Sickle Cell anemia and Focal Segmental Glomerulosclerosis (APOL1 mutation) are not the only situations where a disease mutation protect us against another illness. I’ll just briefly mention two more. Tay-Sachs disease, which is a lethal neurodegenerative disorder in the homozygous state, seems to protect against Tuberculosis in carriers (heterozygotes). Also Cystic Fibrosis (CF) which usually leads to severe and chronic lung disease in the homozygous state, may have protected against the effects of cholera in the heterozygous carriers. The CF mutation inactivates a chloride channel called CFTR, in the cell membrane. Being a carrier for this mutation may have prevented the lethal dysentery that would have accompanied infectious cholera, by preventing water loss in the intestines due to poorly working chloride channels. It is a very common gene mutation, with 1 in 25 people of European descent being a carrier for the CF gene mutation.
When we think disease we think of the suffering of its victims and the cost to society. We are often unaware of the balance of the many forces involved, which influence why a particular disease may be so common in a given population. The factors involved are typically much more complex than we appreciate, and most of them are still unknown to us. Natural selection is working behind the scenes in ways that are difficult to detect on just a casual examination. It may be of no consolation to the sufferers of a serious disease, or the family members devastated by a loved ones sickness and loss, but natural selection, with its cold blind eye to pain or suffering, seems to have fixed some of this in place to allow more genes to be passed onto future generations. Evolution is not directed toward any particular goal and has no empathy or sense of compassion. It only selects those traits that happen to give the organism the best chance to pass on its genes in its evolved environment. This is where the human mind comes into play. Now that we are finally learning to understand the root causes of disease at the genetic and molecular level, we can work to treat, cure, and eradicate disease. Although we are not there yet, in theory it should be possible to cure a condition like sickle cell anemia with gene therapy. At the same time, we shouldn’t have to worry about worsening the burden of malaria if SCA were eliminated, since we can also work on better therapies to treat the malaria, and more effective strategies to prevent infection with Plasmodium.
References and other reading:
1. “Mystery solved: How sickle hemoglobin protects against malaria”, ScienceDaily; April 29, 2011
2. “Sickle Cell Anaemia and Malaria”, Lucio Luzzatoo, Mediterranean Journal of Hematology and Infectious Disease; Oct. 3, 2012.
3. Sickle Cell disease; Wikipedia.
4. Malaria; Wikipedia.
5. Heme Oxygenase-1; Wikipedia.
6. “APOL1 Genetic Variants in Focal Segmental Glomerulosclerosis and HIV-Associated Nephropathy”, Jeffrey B. Kopp, et al., Journal of the American Society of Nephrology; Nov. 2011.
7. “Association of Trypanolytic ApoL1 Variants with Kidney Disease in African-Americans”, Giulio Genovese, et al., Science, August 13, 2010.
8. “A co-evolutionary arms race: trypanosomes shaping the human genome, humans shaping the trypanosome genome”, Paul Capewell, et al., Parasitology, June 26, 2014.
9. “A risk allele for focal segmental glomerulosclerosis in African Americans is located within a region containing APOL1 and MYH9”, Giulio Genovese, et al., Kidney International, Oct. 2010.
10. African Trypanosomiasis; Wikipedia.
Wednesday, July 22, 2015
New Blog site
Darwin's Kidneys has moved to Darwin's Kidneys at Word Press.
Be sure to visit us there!
http://darwinskidneys.wordpress.com
Be sure to visit us there!
http://darwinskidneys.wordpress.com
Can we genetically engineer Rubisco to feed the world?
Today’s atmosphere is brought to you by Rubisco.
Fine makers of oxygen since 3.5 billion B.C.
By Rich Feldenberg

If you happen to peak outside on a nice sunny summers day to admire the green grass, shady trees, and pleasant bushes, your field of view is, in actuality, filled with Rubisco, busily helping the plants do their special thing of making sugar and churning out oxygen. Rubisco is by far the most abundant enzyme on the earth and accounts for 30%, or more of the protein found in the green leaves of plants. Without it there would be no oxygen producing photosynthesis, so if you’re a fan of breathing then you’re probably going to be happy to learn about Rubisco. And, If you’re thinking to yourself, “if there is that much of it in the world it must be doing something important”, congratulations, you’d be right!
There are three things I’d like to point out about Rubisco. One is that Rubisco is freaking amazing. It is an awesome protein with interesting molecular properties, catalyzing fascinating chemistry, and dating back to some of the earliest life on earth. The second thing is that as amazing as Rubisco is, it is incredibly poorly mesigned (mesigned is my word meaning designed by natural selection). Rubisco is horribly inefficient and slow, and it’s amazing it hasn’t been fired from it’s post and replaced with a new, younger, more hip version. And finally, Rubisco could, in principle, be engineered to be much better, possibly increasing crop yields to feed an increasing global population, and removing CO2 from the air to combat global warming. Let's tackle each of these points.
Point One:, Rubisco is amazing. Without it life on earth would likely still be living as simple single celled mats of slime on the ocean floor in a oxygen free world. Rubisco is the abbreviated form for the formal name of the enzyme Rubulose-1,5-bisphosphate carboxylase/oxygenase. Yeah, that’s why Rubisco (rhymes with San Francisco) rolls off the tongue so much nicer, and is way easier to say three times fast. This enzyme goes way back to the good old days when singled cell organisms ruled the world, and appears to have a common origin in all three of the major kingdoms of living things -bacteria, archaea, and eukaryotes- indicating a very early origin sometime around the time of the Last Universal Common Ancestor (LUCA). It appears to have arisen even before the evolution of oxygen producing photosynthesis. There are, by the way, other types of photosynthesis that do not produce oxygen as a byproduct - so called anaerobic photosynthesis- which are less efficient than the oxygen producing types. Based on the study of related proteins, known as Rubisco-like proteins (RLPs), Rubisco may have evolved from RLPs that performed other enzymatic functions before it was eventually modified to it’s modern role in photosynthesis.
Rubisco is a big protein, that is itself composed of two main types of subunits - the large subunit (L) and the small subunit (S). There are three major forms of Rubisco, but in most plants and algae, the Rubisco is composed of a combination of eight L-subunits and eight S-subunits. Rubisco catalyzes the first step in the photosynthetic process taking CO2 and making it react with the compound ribulose 1,5-bisphosphate (RuBP). That is way the Rubisco enzyme is named Rubulose-1,5-bisphosphate carboxylase/oxygenase. In one of it’s reactions, it is carboxylating the substrate RuBP. This leads to the formation of two molecules of phosphoglyceric acid (PGA), that then go through a metabolic pathway called the Calvin cycle. The PGA products eventually go on to other metabolic pathways, and the result is sweet sweet sugar!

Space filling model of the Rubisco protein structure
The L subunit has the active site with a critical lysine residue for binding CO2. It actually takes two CO2 molecules to get things going. The first CO2 molecule is used just as an activator for the enzyme’s active site, but isn’t used in the carboxylation reaction. The second CO2 molecule is what is used to react with RuBP, and it is this carbon that is added onto the molecule. The O2 that is eventually released at the end of photosynthesis does not come from the CO2 but is taken from water, which is also necessary in the reaction. The genes for the L subunit of Rubisco are found in the chloroplasts, tiny organelles within the cells, where Rubisco is conducting its important job. The S subunit is more of a stabilizing part of the protein and its gene is located in the nucleus, and once the protein is made, needs to be shuttled into the chloroplasts.

Also necessary for the enzyme to function is the presence of an ion of Mg+2 ,which acts to stabilize the activation site. This process allows one CO2 molecule, along with a molecule of H2O to become incorporated in RuBP. There is a whole lot of Rubisco in the green leaves of plant to carry out this important chemical reaction. As we said above, about 30% or more of the protein in the leaves of plants is in the form of Rubisco, so it therefore accounts for a huge amount of nitrogen stored in the biosphere, since proteins contain nitrogen as part of their structure.
Point Two: Rubisco is so mind numbingly inefficient I am almost embarrassed for our plant cousins. It turns out that the carboxylase function of Rubisco (you know the really important thing it does by taking CO2 from the air and attaching it to RuBP to begin the process of making carbohydrate) is not the only reaction it performs. In fact, it’s very name -the unabbreviated one that is- tells you right off that it it also is an oxygenase. That is the carboxylase/oxygenase last portion of the name. This means that Rubisco is not terribly selective for CO2, but can also react at the activation site with a molecule of molecular oxygen (O2), which has some chemically similar properties. This leads to a horribly counterproductive metabolic pathway called photorespiration. In other word, it is not very selective, and is so nearsighted that it may grab onto an O2 as easily as a CO2. Normally about 25% of the reactions that Rubisco is catalyzing are with oxygen going down the photorespiration pathway. Also, keep in mind that in the atmosphere today, and for at least the last billion years, the concentration of O2 has been way in excess of that of CO2. The atmosphere these days is 21% O2 and only 0.04% CO2, so that makes it even more difficult for poor little Rubisco to discriminate effectively.

Ribulose 1,5-bisphosphate (RuBP)
Photorespiration leads to RuBP being converted into one molecule of PGA and one molecule of 2-phosphoglycolate. This doesn’t lead to carbohydrate production. Even worse this uses energy in the form of ATP and released CO2 into the air. Totally wrong if you want to store energy from the sun in the form of yummy sugar molecules. So we can clearly say that Rubisco has a poor affinity for CO2. An enzyme’s affinity for its substrate is measured by a characteristic called Km, and Rubisco’s Km is kind of wimpy. This relative non-selectivity may be a reflection of the world in which Rubisco first evolved. At that time the concentration of CO2 in the atmosphere would have been much higher and the concentration of O2 would have been extremely low since photosynthesis was just getting started. Rubisco probably didn’t need to be too selective since O2 was just a trace gas back then.
The selectivity of Rubisco for CO2 over O2 is affected by temperature. Warmer temperatures decrease the selectivity making Rubisco even more inefficient. That can be a problem for plants in a hot dry climate. Also a change in the amounts of CO2 to O2 with respect to each other will influence the enzyme efficiency. These two factors may become a significant concern in a world of global climate change where the both the temperature and concentration of CO2 are on the increase. How this could affect the world’s already insufficient food supplies will have to be seen.
Besides it’s affinity for reacting with a substrate, another characteristic of an enzyme is it’s rate of reaction called the Vmax. Guess what, Rubisco’s Vmax also really sucks. Probably not what you would expect for an enzyme that is the most abundant in the world. Where as most enzymes catalyze thousands of reactions per second, Rubisco is only able to catalyze about 10 reactions per second. Now, I hate to sound so judgmental, but that is really pathetic! It is certainly possible that Rubisco was never able to evolve to be more efficient due to constraints on its structure once it became vital to the plant way of life. Any alteration in the critical active site may have affected too many other protein-protein interactions necessary for normal function, and so never took place. Alternatively, there may be some advantages to photorespiration, after all, so that completely shutting down that pathway would, likewise, be detrimental to growth. There seems to be a trade off between having organisms who’s Rubisco has good affinity for CO2 (Km) and those who’s Rubisco has a fast reaction rate (Vmax). It’s a case of, you can’t have your cake and eat it too. If you favor one quality then you suffer in the other.
Plants have come up with a few smart ways of helping to boost the efficiency of Rubisco. One way is to attempt to concentrate the amount of CO2 around the enzyme. C4 plants do this by adding the carbon from a CO2 molecule to phosphenolpyruvate (PEP), then through a series of chemical reactions, the organic compound malate is produced. The malate is shuttled to the plant cells that contain Rubisco and the CO2 is removed. This concentrates the CO2 in the vicinity of Rubisco so it can act more efficiently. The waste in energy to produce the malate is more than made up for by the better efficiency of the Rubisco in C4 plants due to this CO2 concentrating ability. C4 plants are a more recent evolutionary development, but only represents about 3% of land plants. They are well suited for living in desert conditions where C3 plants would not be able to photosynthesize effectively and would rapidly lose too much water. C3 plants to well in moderate climates with only moderate sun light. The lower temperatures helps to improve Rubisco efficiency at utilizing CO2 over O2.
C4 plants are therefore more efficient, especially in warm dry climates. CAM (Crassulacean acid metabolism) plants close their stomata in the day to prevent fluid loss and open them at night to allow diffusion of CO2 into the leaves, where it is stored in malate. During the day the CO2 is again removed from the malate so it can be used by Rubisco to make carbohydrate. CAM plants can be either land or aquatic.

The C4 plant, Maize, busily concentrating CO2 to boost Rubisco efficiency
Point Three: Perhaps we can make a better Rubisco, one that can select CO2 over O2 more effectively, and react more quickly. Nature has had billions of years to figure this out, so maybe its our turn now to design a Rubisco that can be improved in a variety of different ways. This might be accomplished by artificial selection or genetic engineering - to produce a Genetically modified organism (GMO) with the desirable traits we choose. In fact, there is a great deal of research looking into possible ways to improve Rubisco, but so far progress seems to have been rather modest. A super Rubisco could in theory produce more carbohydrate under warmer, drier, and lower light conditions, decrease the amount of nutrient nitrogen necessary for plant growth, and remove more CO2 from the atmosphere, and release more O2. This could be vital to consider for a growing global population that is outstripping its food resources and heading towards potential disaster due to global warming. How could it be done?
It is known that red algae has a Rubisco with the highest value yet found for CO2 affinity. It is nearly 3 times better at discriminating CO2 from O2 than is the Rubisco from crops, like corn. It may be theoretically possible to engineer crop plants to express the red algae Rubisco. Other studies have looked to genetically engineering Rubisco by substituting key amino acid residues in critical areas of the enzymes protein structure and observing the effect. This has resulted in some mild success. In one study, by switching a particular alanine residue in the L subunit with a asparagine, the affinity was increased by 9%. Not a huge increase, but potentially a good starting point.
Other research has focused on speeding up Rubisco’s slow rate of reaction. One way to accomplish this could be to create a CO2 concentrating mechanism in C3 plants like corn and rice, that is similar to the natural CO2 concentrating mechanisms found in C4 plants. The ways to make this happen are less clear, but could involve manipulations that would put certain types of CO2 transporter in the membranes of chloroplasts to help concentrate the gas where it needs to be.
It should also be noted, that while the intended effects for changes to Rubisco protein would be for the common good of the planet, if we get to the point where such genetically modified plants are possible, it would need to be studied, not only to determine that there are no unintended consequences on the environment, but also that these changes actually result in greater plant growth and yield. There may be some reasons why photorespiration is allowed to occur at the high rates it does. One theory is that this is a protective mechanism for the plant so that in intense light conditions energy overload does not occur that could result in oxidative damage to the plant. There may be a certain limiting factor where carboxylation can be maximized to a certain degree, but once you cross some threshold it actually becomes detrimental to the organism.
There is no doubt that Rubisco is a curious and fascinating protein, and one on which our lives, and continued survival, are completely dependent. It is certainly worthy of our admiration for its important and ancient role in maintaining earths biosphere. There seems to be much more we need to understand about its biochemistry before we can tell if it will be a tool we can utilize to improve and protect our planet. It could also potentially be altered in algae or cyanobacteria to terraform other planets like Mars, which although it has an extremely thin atmosphere, does have an abundance of CO2 over O2. If we eventually discover life on other world that have evolved some form of photosynthesis, it will be interesting to learn what proteins or other methods they came up with to catalyze the carboxylation reaction that Rubisco serves for us here on earth.
References:
1. M. A. J. Parry, et al., “Manipulation of Rubisco: the amount, activity, function, and regulation”. Journal of Experimental Botany, Vol 54, No. 386, pp. 1321-1333, May 2003.
2. Spencer M. Whitney, et. al., “Advancing our understanding and capacity to engineer natures CO2-sequestering enzyme, Rubisco”, Plant Physiology, Vol. 155, pp. 27-35, Jan. 2011.
3. Wikipedia article on Rubisco: https://en.wikipedia.org/wiki/RuBisCO
4. Wikipedia article on Photorespiration: https://en.wikipedia.org/wiki/Photorespiration
5. Article on Mesign: http://darwinskidneys.blogspot.com/2015/07/another-clever-mesign-brought-to-you-by.html
Sunday, July 19, 2015
Origins Sunday: Making nucleic acids may have been child's play on early earth

http://news.sciencemag.org/biology/2015/03/researchers-may-have-solved-origin-life-conundrum
After todays Origin Sunday, I am going to stop doing the short posts and concentrate on the full length weekly articles. Those will go out every Wednesday.
Friday, July 17, 2015
Fossil Friday: Thar She Blows; Whale evolution.
Thar She Blows: Evolution of whales!
by Rich Feldenberg

Welcome back to Fossil Friday. The evolution of the cetacean group (marine mammals like whales and dolphins) is one of the coolest and most beautiful demonstrations of a clear link of fossil evidence from primitive forms to modern forms with many transitional fossils present.

Whales, are of course, mammals and descended from air breathing land vertebrates. All tetrapods descended from lobe finned fish (see my Tiktaalik post). From there they diversified into amphibians, reptiles, mammals, and birds. Some of the groups of reptiles (like the great marine reptiles during the age of the dinosaurs), and mammals (like the whales) returned to the sea many millions of years later. Based on molecular genetics studies, the closest living land mammals to the whales is the hippopotamus.
Cute little Pakicetus was one of the earliest known proto-whales. These hoofed footed mammals were alive about 50 million years ago. Based on bone structure of the skull around the auditory region, they fit into the cetacean group, but were not thought to be good swimmers. Good swimmers in the family would come later!

It is thought that changes in the regulation of genes such as Sonic Hedge Hog (Shh) and Tbx4 may have been important in the loss of the hind limbs in the cetaceans. By affecting when and how genes are expressed, major changes in structure can be made due to relatively small genetic changes. It is also pretty amazing to see the embryology of modern whales also betrays their ancestry. For example, in the whale fetus the nostrils start out in the usual position for a mammal, but as the maxillary bones grow to huge proportions this forces the nasal bones to the top of the skull. This type of evolutionary effect is called allometry and refers to a change in body parts due to changing the growth rate of different parts in relation to one another.
Over time the cetaceans evolved their characteristic echolocation apparatus, as well as, the development the blow hole from nostrils that were originally forward on the face. Today, cetaceans are beautifully adapted for life in the oceans.

References and a cool video to watch:
1. Whale evolution Wikipedia:
2. Animated video of whale evolution. This is pretty cool, check it out.
3. Sonic hedge hog: Wikipedia
Tuesday, July 14, 2015
Why New Horizon's journey to Pluto is so important for us here on earth.
Why New Horizon’s journey to pluto is so important for us here on earth.
by Rich Feldenberg

Almost like a time traveler sent 10 years too far back in time before an important event, I’ve been waiting for July 14th, 2015 for a long time. Ever since the New Horizons space probe was launched from Cape Canaveral, way back on January 19, 2006, I knew this day would get here eventually. It just seemed like our little space probe was taking its sweet time. Nine and a half years is a long time to wait to see a new world - a world never before seen up close and personal. In actuality, New Horizons was doing anything but taking its time. It has been speeding towards its destination at over 36,000 miles/hour! It passed earths moon in a mere 9 hours. It happens to be the fastest man made object ever. It’s just that it had a very long way to go to reach its destination. Today New Horizons will make its closest encounter with Pluto, and almost certainly will increase our knowledge and understanding, not just of Pluto and its entourage of little moons, but of the origins and history of our solar system.
Pluto was only discovered as recently as 1930 by Clyde Tombaugh at the Lowell Observatory. Even from the beginning it seemed a little odd in comparisons to the other planets. It takes about 247 years to orbit the sun and has a very eccentric orbit with its closest point in orbit at 2.7 billion miles from the sun (and inside the orbit of Neptune), and its farthest point in orbit being around 4.5 billion miles away from the sun. It has five known moons, but the largest is Charon, which has a diameter that is more than half as big as the diameter of Pluto itself. No other planet has a moon so close to its own size. For that reason, many planetary scientists consider the Pluto/Charon system a binary system.
Today’s post will go live on Tuesday instead of the usual Darwin’s Kidneys Original Wednesday (sorry Atomic Tuesday) to coincide with this historic occasion. In this post I’m not going to write about the New Horizons discoveries, or much about the mission itself. I’m not even going to write about whether Pluto should be classified as a planet or not. I don’t really care that Pluto got “demoted” to dwarf planet because no matter what we label it, Pluto is a fascinating object with a history as old as our solar system. Instead this article will focus mostly on why we should be interested in a tiny, human made hunk of electronics, computer chips, and metal, speeding to the edge of the solar system to photograph and measure a dark, frozen, ancient celestial body whose chance of harboring life is somewhere between zero and not bloody likely. Why should we, as a society, spend money and resources to design, build, and launch this thing that may not even make it all the way there intact.
We are a species of explorers. Our ancestors traveled the globe and colonized nearly every part of it. We are no strangers to taking risk, and thinking big when it comes to wondering what’s over the next hill or beyond the distant horizon. Human consciousness first awakened on this planet on the continent of Africa, and from there spread to all corners of the world, from stone age Europe and Asia, and over the frozen Bering Straits of the last Ice Age, into North and South America. Early humans even sailed across the forbidding oceans to Australia and the Pacific islands. We have adventure in our blood.

Pluto the most distant target that we have tried, so far, to reach out and touch. Not a journey that humans, with laughably fragile bodies susceptible to harm from radiation and microgravity, and entirely too needy for food, oxygen, warmth, and even companionship, can make anytime soon. Instead we send our stoic little robot ambassador out on a entirely peaceful mission of scientific discovery. It represents the best part of humanity with no thought whatsoever to invasion, conquest, or exploitation of new territory for gain or profit. It represents what’s best in us - our childlike curiosity, enthusiasm for discovery, and sense of awe at living in a universe that is so much bigger than our everyday concerns.
Going to Pluto inspires us to be great by doing great things. It is no trivial task to design, build, and implement a machine to do what New Horizons is doing right now. That’s, of course, why it has never been done before. The accuracy necessary for the mission to reach its target, and the durability of its components to remain functional after 9 years in the cold vacuum of space, are a triumph of human engineering and understanding of Newtonian mechanics. The challenge of the mission elevates us up onto a more noble plane. Teams of individuals made the mission possible, but also the millions of taxpayers that contributed to a successful human achievement, are all part of the process that show we as a society care about things beyond the mundane and everyday. We are all apart of the mission, and we all have a right to see what New Horizons can tell us about the edge of our cosmic neighborhood block.
Going to Pluto also inspires curiosity in the unknown. From earth, even from the Hubble Space Telescope, Pluto is not much more than a dim dot in the night sky. We want to know, what is it like on Pluto? Why is it so different than the planets like the Earth, Mars, Jupiter, and so on? What is it made of and why is its orbit around the sun so unusual? Does it hold clues to the formation of the solar system and the planets? Could it hold clues to the origins of life’s chemical building blocks that lead to our own origins on earth? We humans really want to know the answers to things. When we have a real mystery it inspires a lot of careful thinking, formulation of hypotheses, and ideas about how to test those hypotheses. Being curious is one of our most outstanding traits as a species. Far from the old adage “curiosity killed the cat” in reality, curiosity is how we learn who we are, where we come from, and what our place in the universe really is. “Curiosity killed the cat” is meant to keep us afraid and in the dark. Curiosity keeps us moving forward, but the spirit of curiosity is easily doused by others who are perfectly satisfied by not knowing and who have long ago lost their curiosity. We need to keep that spark of curiosity alive. Not only is a mission like New Horizons the scratch to satisfy the itch of our innate curiosity, but it inspires new levels of curiosity in those sharing in the discoveries, and in the imaginations of young people who then begin thinking about what is next out there to explore.

There are also the unforeseen consequences from a mission like New Horizons. It is not why these missions are undertaken, but we have reaped the benefits of the collateral developments (the opposite of collateral damage) of basic science research before. From the World Wide Web developed by theoretical physicists at CERN, to advances in computer and laser technology, basic science research has provided benefits to society at large that were never predicted or expected. When Nobel prize winning physicist, Edward Purcell was asked what practical applications his discovery of nuclear magnetic resonance in bulk matter could ever be used for, which he developed to better understand the quantum transition of hydrogen atoms from one energy state to another, his answer was, “I can see no practical applications”. It turned out that this discovery changed modern life giving us Magnetic Resonance Imaging (MRI) in medicine to peer into the living body in exquisite detail, as well as transforming the field of chemistry with Nuclear Magnetic Resonance (NMR) which has revolutionized our understanding of molecular structure and material science. The truth is that we don’t always know what the final impact of fundamental research may be for our everyday lives. The knowledge we gain from studying Pluto might help us better understand the threat of comets and asteroids to life on planet earth, and perhaps aid in our survival as a species. The most likely benefit will be ones we don’t see coming at all. There are also economic gains that programs, such as the space program provide to our country, as far as more jobs, and it signals to the world our national strengths and that intellectual endeavors are an important priority. Being a leader in science and space exploration is no small thing in the eyes of the rest of the world.
Missions like New Horizons remind us that we live in a much bigger universe than we are used to thinking about. On a day to day basis, it’s easy to focus on the minor details, to think your little neighborhood is all there is. We don’t look up at the night sky and observe the stars very often- not nearly enough. Going to Pluto forces us to think about our place in the cosmos. The solar system is big and the planets are far away. How much bigger is our galaxy than the solar system, and what about the billions of distant galaxies? We are not just in the universe, the universe is in us. We are a part of the universe and it’s good to be reminded of that from time to time.
I’ve waited a long time for today. I don’t know what pictures and information will be sent back to earth by our little robotic probe as it speeds past Pluto, but I know it will be amazing. Just to know that something of earth is out there, so far from home and continuing its flight outward into the galaxy, is pretty cool in itself. And once New Horizons leaves Pluto behind, there will continue to be new and exciting discoveries to anticipate, some from future robotic space missions, others from telescopic observatories examining the universes largest structures, and still others from basic science research facilities like CERN examining the universes smallest components and fundamental forces. We will continue to have a lot to learn and look forward to so long as we as a society continue to decide that the nobel pursuit of new knowledge is a goal worth achieving. For today, I just want to say, “Hello Pluto, it’s great to finally meet you”.
References:
1. NASA New Horizons website.
2. Pluto: Wikipedia
3. Cylde Tombaugh: Wikipedia
4. Pluto Safari is a cool app you can down load on your tablet from iTunes.
Subscribe to:
Comments (Atom)